
��(d��ng)ǰλ�� > ��� > ���g(sh��)���� > ��˥�ϳ��w�S������(j��ng)���⾀�p�����DNA �ޏ�(f��)�ͼ��������{(di��o)��
��˥�ϳ��w�S������(j��ng)���⾀�p�����DNA �ޏ�(f��)�ͼ��������{(di��o)��
��˥�ϳ��w�S������(j��ng)���⾀�p�����DNA �ޏ�(f��)�ͼ��������{(di��o)��
�ν����������� ̹ͯ��
(�����t(y��)�ƴ�W(xu��)���ﻯ�W(xu��)�c��������W(xu��)ϵ, ������100083)
ժҪ�����w�����B(y��ng)�IJ�ͬ���g�����߷ζ����w���w�S����(2BS) �錦��, ���⾀�T��(d��o)DNA �p����, �^�켚���ΑB(t��i)����ֳ�������������ڡ�DNA�ޏ�(f��)׃���ȼ�����(y��ng)���Լ�gadd153��p21��p53 �Ȼ�����D(zhu��n)�ˮƽ�ı��_׃��. �Y(ji��)���@ʾ: ���⾀�T��(d��o)DNA�p����, ˥��(>55��) 2BS ������
�B(t��i)����ֳ�����ĸ�׃�������p����(<30��) �@��; ��ͬ���g�ļ����p��������F(xi��n)G1 ������F(xi��n)��, ���p����G1 ����������@����˥�ϼ���(P<0105) ; ˥�ϼ��������ޏ�(f��)�����^���p�������@�½�(P<0101) ; ͬ�r, gadd153��p21��p53 �ȵĿ��T��(d��o)�Ծ��������p2BS ����. �ɴ�, �քe�ڼ���ˮƽ�c����ˮƽ��ӳ��˥�ϼ�����(j��ng)���⾀����p����ļ�����(y��ng)��׃���c�ޏ�(f��)�C�ܜp�˵��P(gu��n)ϵ.
�P(gu��n)�I�~��˥�ϼ���,DNA �p���ޏ�(f��), ��������, �z��c����
��DNA �p���ķe�ۼ��ޏ�(f��)�������½�������˥�ϵ���Ҫԭ��֮һ[1]. p53��p21 ��ATM �ǽ���l(f��)�F(xi��n)�������3��DNA �p����(y��ng)��ęz��c����(check2point control) ����[2]. p53��p21 ���H�����ְ�����l(f��)�]��Ҫ����, ����DNA �p���ļ�����(y��ng)�����������{(di��o)������, �������{(di��o)��(ji��)DNA�p���ޏ�(f��)��������������Լ������������T�������D(zhu��n)�[3]. DNA�p�����T��(d��o)�����е�gadd45��gadd153�cDNA �p����ļ�����������Լ��ޏ�(f��)Ҳ�������P(gu��n), ��������p53�����λ����������[3]. �@Щ������˥�ϼ������Пo���_׃�����c˥�ϼ����ޏ�(f��)�������½��Ƿ����P(gu��n)? ����δҊ����(n��i)����. �҂����w�����B(y��ng)�����߷ζ����w���w�S����(2BS)�錦��, �о������⾀�����˥�ϼ����е�DNA �p�����T��(d��o)����gadd153 �Լ��z��c���ƻ���p21��p53�ı��_׃��,�����cDNA�ޏ�(f��)�����ͼ�������׃�������P(gu��n)��.
1�������c����
1.1�����ϼ���Ҫԇ�������߷ζ����w���w�S���������l(w��i)����������Ʒ�о���, pB luesSK �|(zh��)��(gadd153 cDNA ) �ɱ����t(y��)�ƴ�W(xu��)�����t(y��)Ժ��ٛ,p21WAF1.CIP1. SDI1cDNA������Baylor �t(y��)�W(xu��)Ժ��ٛ,p53��B2act in cDNA ̽ᘞ鱾������. DMEM�ɷ����B(y��ng)��: GIBCO.BRL��˾, ̥ţѪ��: ���������r(n��ng)��ѪҺ��Ʒ��, 3H2TdR ��A232P dCTP: �������x���﹤�̹�˾, P rim e2a2Gene Lebeling System:Promege��˾, �u�������q�~��DNA������ϩ������ͪ��ţѪ�����: Sigma��˾, ����ԇ������Sig2ma�a(ch��n)Ʒ����a(ch��n)������.
1.2���������B(y��ng)�����߷ζ����w���w�S����(2BS)���B(y��ng)�ں�10% ̥ţѪ���DMEM ���B(y��ng)Һ��, 37������������L, ������62��4 ����24��6 ���քe����˥�ϼ����p����ʹ��.
1.3�������ΑB(t��i)����ֳ�����^�졡�������L������(sh��)���L�ں�, ������Һ�{(di��o)����1��104����.ml, ����24�װ������B(y��ng), �������N�ں������⾀(0145J.m 2.s) ����5 min �p������[4],���Q���r���B(y��ng)Һ��37�����, ÿ12h�^�켚���ΑB(t��i)׃�����M�м���Ӌ��(sh��), ��δ�M�����⾀�����˥�ϼ����p����������.
1.4���������ڷ��������⾀���䄩�����r�gͬ��,37�����24h����ø����, �ռ�����, ����RNA ø��37������1h, Ȼ���õ⻯���(PI)Ⱦɫ, ��Bec2ton2Dickinson ��ʽ�����x�M��FACScan(fluo res2cence act ivated cell sorting)����, ��δ�M�����⾀�����˥�ϼ����p������������
1.5��UDS (un scheduled DNA syn thes is) �y��������Ӌ��(sh��)��, ��1��105 ����.�ׂ���6�װ�, ��������ȫ�N�ں�,�Ժ�015% ̥ţѪ���DMEM���B(y��ng)Һ���I����72hʹ����ͬ����,���Q��5mmo l.L�u����ğoѪ��DMEM ���B(y��ng)Һ��37�����1h��,���⾀����, �䄩���c�r�gͬ��,���Q���B(y��ng)Һ(��5mmo l.L �u���塢1LCi.m l3H2TdR��10%̥ţѪ���DMEM) , 37�������ͬ�r�g�ռ�����,�M��3H2TdRӋ��(sh��), ���սM��δ�M�����⾀����ļ���[5]��
1.6�����c��Northern ӡ�E�s������̎�ڌ���(sh��)���L�ڕr�M�����⾀����,�r�g������ͬ��, Ȼ����37�������ͬ�r�g�ռ�����, �î���������һ������ȡ������RNA,Ȼ���D(zhu��n)Ĥ����(bi��o)ӛ̽ᘼ��M�а��c��Northern �s��[6,7] , ��δ�M�����⾀����ļ����錦��. �s���Y(ji��)���ɷe�ֹ��ܶȒ��������
2���Y(ji��)��
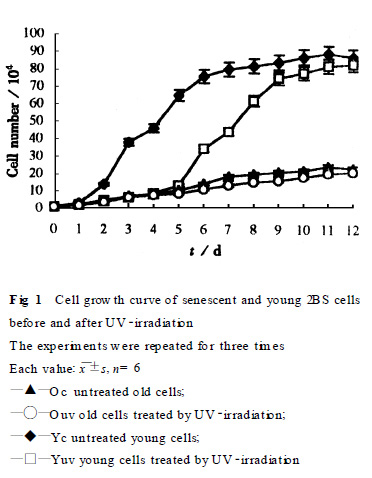
2.1�������ΑB(t��i)����ֳ���Եı��^�����⾀����ǰ��Ҋ˥��2BS�����ΑB(t��i)�c���p�������@��ͬ, ˥�ϼ������w�ʴ������{��(n��i)�w�����ࡢ���{������۹����½����������ИO��Ҏ(gu��)�t; �����p�����t���w���L�����{���������������u������. ���⾀�����, ˥�ϼ����ΑB(t��i)׃����ʮ�����@, �HҊ���{��(n��i)�w�����@����; �����p�����t��Ҋ���{�w�����@���ࡢ���{�����½������wҲ���@����, �����B(y��ng)�s1�ܺ��ΑB(t��i)�����֏�(f��)����. ���⾀����ǰ�����L����ҊFig.1, ��(j��ng)���⾀��������p������ֳ�����ܵ����@����, ���s6d����ֳ�����֏�(f��)����; ˥�ϼ�����ֳ�������@�������p����, ���⾀����ǰ����ֳ׃����ʮ�����@��
212���������ڕr��׃�������⾀�����˥�ϼ����p��2BS ��������ҊG1 �����, �����p����G1 �ڼ�������ʸ����^˥�ϼ���(P<0105) (Fig.2).

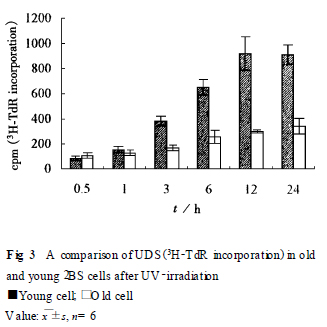
213 DNA �ޏ�(f��)�������^����3H2TdR ���뷨�y���dz�����DNA �ϳ�(UDS) , �Ա�ʾDNA ���ޏ�(f��)����. ��F ig. 3 ��Ҋ, �����⾀�����12 h ����UDS�_��ֵ. ˥��2BS �������ޏ�(f��)�������@�������p����(P < 0101).
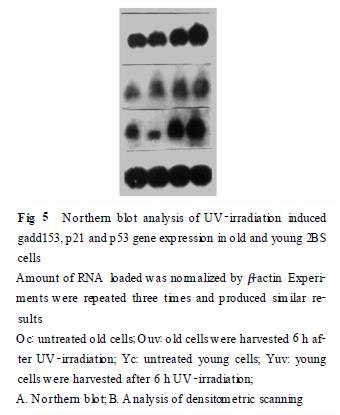
214�����c��Northern ӡ�E�s���Y(ji��)���������ɰ��c�s���Y(ji��)����Ҋ, ���⾀����6 h ��gadd 153 ���_�_��ֵ, ˥�ϼ�����gadd 153 ���T��(d��o)�����@�������p����, �Y(ji��)��Ҋ(Fig. 4). ����(j��)���c�s���Y(ji��)���xȡ���⾀�����6 h �ռ�����, �M��Northern �s��, �Y(ji��)���@ʾ(Fig. 5) , ���⾀�����˥�ϼ�����g(sh��)add153��p 53��p 21 ����Ŀ��T��(d��o)�Ծ����@�������p����, ������p 53 �T��(d��o)�Ը�׃��С. ����, �@���N���������⾀�p��ǰ�Ļ������_��˥�ϼ��������p������Ҳ������ͬ, ����p 21 ��˥�ϼ����еĻ������_���@�������p����, gadd153 �ı��_��˥�ϼ��������Ը������p����, ��p53 �ı��_����@��.

3��ӑՓ
����˥����һ������������;�����µď�(f��)�s�Y(ji��)��,DNA �p���ķe�ۼ��ޏ�(f��)�������½������Ǽ���˥�ϵ���Ҫԭ��֮һ[8] , ��DNA �ܓp��˥�ϼ�����(y��ng)���c���p����������ͬ. ˥�ϼ������ΑB(t��i)����ֳ�����ĸ�׃�����@, ͬ�r��֏�(f��)Ҳ����. ��ʽ����Ӌ�z�y�Y(ji��)���@ʾ, ˥�ϼ���G1 ������ʵ����ӷ��ȵ������p����; ͬ�r˥�ϼ�����DNA �ޏ�(f��)�����@���������p����. �@Щ�Y(ji��)�����f��˥�ϼ�������DNA �p���đ�(y��ng)���������@�������p����. ��ô, ˥�ϼ�����DNA �p���đ�(y��ng)�������½��ĸ���ԭ����ʲô��? p53��p21 ��gaddl53 �Ȼ�����_��r�����⾀����ǰ���׃�����ܞ��҂��ṩ��һЩ�Ѓrֵ�ľ���.
DNA �p����, ��ͨ�^һ������̖����(d��o);����������(y��ng), �T��(d��o)���������_�ļ��������,DNA �ޏ�(f��)ϵ�y(t��ng)�ļ���t�Ǽ����S���������������Ժͼ��������һ����Ҫ�C��[ 9 ]. p 53 ��DNA�p���O(ji��n)����̎�ڙz��c���Ƶ�λ��, �����И�����,p21 ��gadd153 ���������λ���, �����������ֹ�ͬ��;��, �ֿ�ͨ�^���Բ�ͬ��;�������ڼ������ڼ��ޏ�(f��)ϵ�y(t��ng), ��DNA �p���ޏ�(f��)�����{(di��o)�ؾW(w��ng)�j(lu��)�мȻ����f(xi��)ͬ���Ӱ�, �l(f��)�]�����Բ�ͬ������[9]. p21�����ڵ���ø����������,����p 53�����λ���, ��p 53�c���������{(di��o)��֮�g���������. p 21�ĸ߱��_����(d��o)�¼����������, ߀��ͨ�^�cPCNA (��ֳ�����˿�ԭ) ������Ӱ�DNA �ď�(f��)�Ƽ����ޏ�(f��)[3]. gadd153 �����⾀���T��(d��o)���_��Ҳ����ه��p53��, ������p53 �����λ���,һ���挦�p����ļ���������һ��Ӱ�, ��һ��������cDNA �p���ޏ�(f��)ż(li��n). ���īI���, gadd153���D(zhu��n)�����C/EBP (CAAT�����ӽY(ji��)�ϵ���) ������ؓ�{(di��o)������[10]. ���Y(ji��)���@ʾ, ��(j��ng)���⾀�p����oՓ���p��˥�ϼ������F(xi��n)��G1 �ڵ����ͬ�r���S��gadd153��p 21��p 53 ����ĸ߱��_, �@�f���@���N����Ŀ��T��(d��o)�c���������������ֱ�ӵ��P(gu��n)ϵ; Ȼ��, ��Ȥ����˥�ϼ��������⾀����ǰ���������@��G1 �����, �˕r�HҊp 21 �ĸ߱��_, �@�f��p 21��˥�ϼ���G1 �ڵ������������Ҫ����, ��p21�ĸ߱��_�����˥�ϼ����д��ڵēp���۷e���ܱ���Ч�ޏ�(f��)����. ���˥�ϼ����������������@��G1�����, ��֮p 53��gadd153, ������p 21 ������T��(d��o)���½�, ������˥�ϼ���G1 �������������p������ԭ��. ͬ�r, ˥�ϼ����ޏ�(f��)�������½�Ҳ�����cgadd ����(����gadd153, gadd45 ��)��p 21 ��p 53 �Ȼ����DNA �p����Ŀ��T��(d��o)����˥�ϼ����е��½����P(gu��n). ���Y(ji��)���@ʾ, ˥�ϼ������mȻp 21 �@һ���������������ӵĸ߱��_��(d��o)���˼�����G1 �����, ���ޏ�(f��)�p����ȡ�˕r�g, �������c�ޏ�(f��)ֱ�����P(gu��n)�Ļ���s������Ч�ر��_�Ͱl(f��)������. �����߀�l(f��)�F(xi��n), p 53 ���_׃������ʮ���@��, �f��p 21 ��˥�ϼ����еĸ߱��_�п���ͨ�^��p 53 ;���{(di��o)��[ 10 ]; ����һ�����@�����cp 53 ����ͨ�^���ữ��B(t��i)�ĸ�׃�����F(xi��n)���P(gu��n)[ 11 ] , �ړp���T��(d��o)��, p 53 �����c���N����ĽY(ji��)�ϻ��Կ���ͨ�^�����ữ��B(t��i)�ĸ�׃��������׃, �@ͬ�ӿ�����˥�ϼ�����p 53 δҊ�@���߱��_��ԭ��֮һ. Ȼ��, �@�N������˥�ϼ����еēp�����T��(d��o)���½��s����һ����, �@��������˥�ϼ�����DNA �p���������{(di��o)���������ޏ�(f��)�����½��ĸ���ԭ��֮һ. ��֮, ̽ӑ˥�ϼ�����DNA �p���������{(di��o)����DNA�p���ޏ�(f��)�Լ�DNA �p�����T��(d��o)����ı��_��׃, ������DNA �p���ޏ�(f��)�����{(di��o)�ؾW(w��ng)�j(lu��)���U��, ���ڽ�ʾDNA �p���ޏ�(f��)�c����˥�ϵ��P(gu��n)ϵ������Ҫ����Փ���x.
References
1 ��̹ͯ��, ������. �t(y��)�W(xu��)����W(xu��)����˥���c�L��. ����: �����l(w��i)��������, (Tong Tanjun, Zhang Zongyu. M ed ical Gerontology ����Senescence and long ev ity. Beijing: People��s Health P ress) 1995:139�� 142
2 ��Paulovich A G, Toczysk iD P, Hartwell L H. When checkpoints fail. Cell, 1997, 88: 315��321
3 ��Scanchez Y, Elledge S J. Stopped fo r repairs. B ioE ssay s, 1995, 17(6) : 545�� 548
4 ��������, ������, �S��, �����}. ˥���^���д���Ƣ����DNA �ޏ�(f��)������׃��, ���A�����t(y��)�W(xu��)�s־, (N an Xinsheng, ZhangZongyu,Huang L i, Zhang Changying. Change of DNA repair ca2pacity of rat sp leen cells during aging. Ch in J Geriatrics) 1992,11 (5): 301
5 ��T illey R,M iller S, Srivastava V , Busbee D. Enhanced unsched2uled DNA synthesis by secondary cultures of lung cells estab2lished from calo rically restricted aged rats. Mech Ageing D ev ,1992, 63: 165�� 176
6 �����Ĺ�, ̹ͯ��. ���������^����c2erbB22 ����ı��_. �Ї����ﻯ�W(xu��)�c��������W(xu��)��(W angW engong, Tong Tanjun. Theex2pression of c2erbB22 on apoptotic process. Ch in J B iochem M olB iol) , 1998, 14 (3) : 314�� 317
7 �����Ĺ�, ͯԫ��. С����w�S���������^����p53 �cbcl22 ���_�ĕr����. �Ї����ﻯ�W(xu��)�c��������W(xu��)��(W ang W engong,Tong Tanjun. The time o rder of p53 and bcl22 gene exp ressionin the apop to tic p rocess of mouse fibroblast cell lines. Ch in JB iochem
M ol B iol) , 1998, 14 (3) : 318�� 321
8 ��Vo jta P J , Barrett J C. Genetic analysis of cellular senescence.B ioch im B iop hy c A cta, 1995, 1242: 29�� 41
9 ��Sancar A. DNA repair in humans. A nnu R ev Genetics, 1995, 29:69��105
10��Delmastro D A , L i J , V aisman A , So lle M , Chaney S G. DNAdamage inducible gene exp ression fo llow ing p latinum treatmentin human ovarian carcinoma cell lines. Cancer Chem other P har2m acol, 1997, 39: 245��253
11��V aziri H,W estM D,A llwopp R C, Davison T S,W u Y S, A r2row smith C H, Poirier G G, Bench imo l S. A TM 2dependent telomere loss in aging human dip lo id fibroblasts and DNA dam2age lead to the po st2translational activation of p53 protein in2vo lving po ly (ADP2ribose) polymerase. EM BO J , 1997, 16 (19) :6018��6033

- �Ї��t(y��)�W(xu��)�b������2025�t(y��)�W(xu��)�b��չ�[��֪ͨ
- analytica China 2022����O(sh��)������㲿��չ�^(q��)����
- 2022�������������c����ҽ��O(sh��)�������[�����ڹ���
- ���i�Ƽ�������������x����У��(zh��n)Ҏ(gu��)��
- ��̩�˿�Ҋ����x/���z�x���Mԇ����
- �Ϻ����i�\�������^�Ͼ��ƌW(xu��)�x����������b��չ
- �Ϻ����ʹ�˾���Ƴ������w����b�á�
- ��ʮ�Č�ȫ���R���[���W(xu��)����� CSCO �W(xu��)�g(sh��)���֪ͨ


Copyright(C) 1998-2025 �������ľW(w��ng) �Ԓ��021-64166852;13621656896 E-mail��[email protected]